Huntington’s disease (HD) is a monogenetic disorder with a spectrum of clinical features. Besides the characteristic involuntary motor disturbances, such as chorea, individuals affected by the disease suffer from psychiatric symptoms and cognitive decline.1–3 Despite this, focus on the movement disorder in clinical practice and the research of HD has been strong.
Huntington’s disease (HD) is a monogenetic disorder with a spectrum of clinical features. Besides the characteristic involuntary motor disturbances, such as chorea, individuals affected by the disease suffer from psychiatric symptoms and cognitive decline.1–3 Despite this, focus on the movement disorder in clinical practice and the research of HD has been strong.
In recent years, studies of non-motor symptoms and signs in HD have revealed that sleep disturbances, disruption of the circadian rhythm, autonomic dysfunction and metabolic alterations are also common and occur early on in the disease process. Therefore the conventional notion that HD is solely a movement disorder caused by selective basal ganglia pathology had to be challenged.
The fact that the hypothalamus and its neuroendocrine circuitries are important regulators of emotion, sleep and bodyweight has stimulated research investigating to what extent this system is affected in HD. Although further studies are needed, the current literature now provides strong support for hypothalamic pathology and neuroendocrine alterations to be included as part of the clinical phenotype of HD.
This article reviews the current state of knowledge in terms of the clinical relevance of hypothalamic and neuroendocrine changes in HD.
The Genetics of Huntington’s Disease
This hereditary and monogenetic neurodegenerative disease has a prevalence of around 1:10,000.1 It is caused by an expanded CAG repeat in the huntingtin gene, which codes for an expanded polyglutamine in the huntingtin protein.4 This protein is expressed in all tissues in the body. Although huntingtin protein’s normal function is not fully known, it is thought to involve vesicle transport, transcriptional regulation and synaptic function.5,6 CAG repeats of 40 or more cause the disease with full penetrance, with onset of motor symptoms in midlife. Huntingtin genes with between 37 and 39 CAG repeats have reduced penetrance. Intermediate repeats between 29 and 35 do not cause the disease but may expand to a pathogenic range in future generations. Juvenile onset (<20 years of age) occurs with more than 60 to 70 CAG repeats. Hence, longer CAG repeats are associated with a younger age of onset. The CAG repeat length accounts for around 60% of the variance in age of onset; genetic and/or environmental factors are thought to cause the rest.7 Interestingly, the genetic modifiers that have so far been reported to influence the age of onset in HD include factors involved in the regulation of metabolism, such as huntingtin-associated protein 1 (HAP1) and PPARγ coactivator-1α (PGC-1α).8,9
Predictive and diagnostic genetic testing for HD is now available. Despite this, only around 15% of all individuals who develop the disease choose to undergo predictive testing, i.e. genetic testing, before they manifest any symptoms.10
Clinical Features
Clinical diagnosis currently requires unequivocal signs of motor disturbances, such as chorea, and death usually occurs 15 to 25 years after the onset of motor symptoms.1,2 Psychiatric symptoms are common and include depression, anxiety and irritability.11–13 There is an increased prevalence of suicidal ideation and suicide attempts in HD, particularly after receiving the formal diagnosis and at a later stage when independence diminishes.14 Early cognitive changes include a ‘frontal behaviour’ characterised by apathy, disinhibition and executive dysfunction.15 Other cognitive changes involve reduced processing speed and attention, and often progress to dementia.16 Psychiatric and cognitive changes are now thought to precede the motor symptoms by many years.17 Other non-motor features are also common and occur early. Sleep disturbances have a prevalence of around 90% and are characterised by an increased latency of sleep onset, reduced sleep efficiency, frequent nocturnal awakenings and delayed and shorted rapid eye movements.18–21 HD patients also have an altered circadian rhythm and autonomic dysfunction.22–25 Metabolic changes in HD include increased appetite, increased metabolism and weight loss in advanced stages of the disease.26–28 In summary, the clinical presentation of HD is characterised by a relatively wide range of non-motor features that often precede the progressive motor symptoms. Therefore, unraveling the pathogenic mechanisms and the neurobiological origin of these early non-motor features is therefore likely to be crucial in understanding the full disease process and in identifying novel targets for disease-modifying intervention.
Pathology in the Basal Ganglia and Beyond
Cell loss and gliosis in the striatum of the basal ganglia are hallmarks of HD.29 Medium spiny-aminobutyric acid (GABA)-ergic neurons in the striatum are selectively affected; whereas NADPH-diaphorase-expressing interneurons are spared.29 The staging of neuropathological changes in HD is based on the extent of these changes in the striatum.30 Loss of neurons and general atrophy also occur in the cerebral cortex and have recently been studied using imaging techniques.31–33 Neuronal intranuclear inclusions (NIIs) of aggregated mutant huntingtin are present in these regions in HD patients.34
Interestingly, a study of a transgenic HD mouse model revealed that NIIs were present in many tissues in the body.35 These results drew attention to the possibility that mutant huntingtin may exert pathological effects beyond the basal ganglia and the cerebral cortex. Indeed, peripheral tissues in HD patients display several pathological alterations. These include skeletal muscle wasting with mitochondrial dysfunction, atrophy of the testis with reduced testosterone production and hyperactive monocytes with peripheral immune activation (recently reviewed in van der Burg et al.).36 The notion of HD being a selective basal ganglia disorder has hence been profoundly challenged.
Neuropathological Changes in the Hypothalamus in Huntington’s Disease
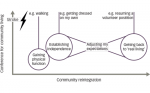
The hypothalamus consists of a number of interconnected nuclei that express a variety of hormones and neuropeptides involved in control of the endocrine system as well as in the regulation of emotion, metabolism and sleep.37–42 Studies investigating alterations of the hypothalamus and the neuroendocrine system in HD began over 60 years ago. The findings so far in both clinical HD and in animal models of the disease have been discussed in detail in a few reviews published in recent years.43–46 The major positive results from studies investigating alterations in the hypothalamic and neuroendocrine system in clinical HD are illustrated in Figure 1.
Few studies have focused on neuropathology in the hypothalamus in HD. The first systematic analysis of a hypothalamic nucleus in HD was performed by Kremer et al., who described loss of somatostatin neurons in and atrophy of the nucleus tuberalis lateralis.47–49 The function of this specific nucleus is still unclear. Loss of the neuropeptide orexin (also called hypocretin) – implicated in the sleep disorder narcolepsy as well as in the control of emotion and glucose metabolism – was demonstrated in the lateral hypothalamus of HD patients.50,51 Although the loss of around 30% of orexin-expressing neurons in HD was not sufficient to be reflected in the cerebrospinal fluid (CSF), this finding inspired further studies focusing on sleep in HD as well as on the full extent of neuropathological changes in the hypothalamus in HD.52–55
Progress of the investigations into hypothalamic changes in HD has been limited by the scarcity of tissue from this region in brain banks and due to lack of a clear morphological definition of the borders of this region and its nuclei. A method has recently been established to accurately and in a reproducible fashion delineate this region using robust anatomical landmarks in brain sections stained with the cell marker cresyl violet and the myelin-staining luxol fast blue.56 This technique has been used to perform a systematic stereological analysis of the hypothalamic region in HD. A trend towards atrophy of the whole hypothalamic volume was found in a relatively small cohort of nine HD cases compared with eight controls.56
Analysis of specific nuclei, such as the paraventricular nucleus, revealed that there was significant loss of neurons in this area in HD cases compared with controls.56 Interestingly, reductions were also detected in the number of oxytocin- and vasopressin-expressing neurons in the HD hypothalamus. These neuropeptides have both been implicated in social behaviour.38 In particular, oxytocin has been found to increase trust, empathy and facial memory.38,57,58 Therefore, it is possible that loss of oxytocin could be involved in the reduced recognition of facial expression of emotions that has been reported in HD patients.59–65 Neuronal populations expressing neuropeptide Y and melanin-concentrating hormone were not found to be altered in the HD hypothalamus.50,56 Hence, not all neuropeptide-expressing populations in the hypothalamus are affected in HD. Taken together, the neuropathological studies to date show that there is neuronal loss in specific nuclei of the human HD hypothalamus as well as alterations in emotion- and metabolism-controlling neuropeptides.
Imaging of the Hypothalamus in Huntington’s Disease
The hypothalamic region has no clear anatomical borders and therefore provides a challenge for delineation in magnetic resonance images (MRIs). Using voxel-based morphometry (VBM), significant differences have been found in the grey-matter contents in the hypothalamic region between HD patients with motor symptoms and age- and sex-matched controls.66,67
A recent study continued to investigate the extent of changes in the hypothalamic region using MRI scans from the PREDICT-HD study. The PREDICT-HD study is a multicentre observational study that has enrolled a large number of individuals who have tested positive for the disease-causing huntingtin gene but who do not yet have motor symptoms, i.e. prodromal HD.68 A significant reduction in gray matter signal was found in the hypothalamic region in prodromal HD that paralleled alterations in the striatum and insula over a decade before the expected onset of motor symptoms using VBM.69
A different approach based on mathematical modelling revealed that the gray matter content alone in the hypothalamic region in these MRIs was powerful enough to distinguish prodromal HD from controls. It was also possible to distinguish different groups of prodromal HD divided on expected time to motor onset using this approach. Another study using positron emission tomography has found reductions in dopamine D2 receptors as well as microglia activation in the hypothalamic region of prodromal HD.70 Taken together, these results show that the hypothalamic region is affected early on in the pathogenesis of HD.
Alterations of Neuroendocrine Factors in the Cerebrospinal Fluid, Blood and Urine
It is well-known that some factors secreted from neurons in the central nervous system can be measured in the CSF. In other neurodegenerative disorders, such as Alzheimer’s disease, alterations in CSF levels of tau, phosphorylated tau and the 42 amino acid form of amyloid-beta proteins are considered to reflect the disease process.71 Measurements of these factors today guide the diagnosis of Alzheimer’s disease and are used as biomarkers.
Biomarkers are usually divided into trait biomarkers – indicating a specific disease – and state biomarkers—reflecting the state in the disease process. In HD, the trait marker consisting of the mutant huntingtin gene is well established. State markers, which reflect disease process before the onset of motor symptoms in HD, are urgently needed for the evaluation of new, potential treatments. As factors in the neuroendocrine system can be measured in CSF and serum, and given the fact that this system is affected in HD, the hope of finding an altered neuroendocrine factor as a state biomarker for the disease has emerged. However, the search for a neuroendrocrine factor suitable as a biomarker is complicated. Such factors are part of interconnected and complex circuitries where the overall system strives to maintain homeostasis. Therefore, each factor is closely regulated. This is not the only challenge – a change in the level of one factor may be primary, due to the effects of the mutant HD gene, or secondary, due to changes in another part of the circuitry. Neuroendocrine factors may also be affected by the diurnal rhythm, satiety level, gender, age, medication and so forth. Variation in the available assays used to measure neuroendocrine factors is another major consideration. The reports published so far investigating the neuroendocrine system using CSF and blood measurements in HD have suffered from a large variation between and within studies.43–46
A few clinical studies investigating neuroendocrine factors in HD have revealed promising results. Increased levels of cocaine and amphetamine-regulated transcript (CART) have been found in the CSF. These levels may reflect the increased numbers of CART-immunopositive neurons detected in the human HD hypothalamus using immunohistochemistry.56,72 CART is a neuropeptide known to increase anxiety-like behaviour in animal models and has been implicated in mood disorders.73–75
Analyses of the hypothalamic–pituitary–adrenal (HPA) axis in blood and urine have collectively pointed to an upregulation of this endocrine axis in HD.76–78 An activated HPA axis has been one of the most consitant findings in clinical depression and may have negative effects on cognitive function and energy homeostasis.79 Moreover, a delayed onset in the diurnal rise of melatonin has been found in HD patients, suggesting dysfunction in the hypothalamic suprachiasmatic nucleus.80 This nucleus functions as a circadian clock, being the principal rhythm-generating system in humans.
Changes in hypothalamic function can also be reflected in alterations in afferent signals from the periphery. These afferent signals to the hypothalamus include:81
• leptin, a satiety signal from adipose tissue;
• ghrelin, an appetite signal from the gastric mucosa; and
• insulin, an anabolic peptide secreted from the pancreas.
Indeed, insulin resistance has been found in the early stages of HD.82 Reduced leptin and increased ghrelin levels have been reported in later stages of HD, possibly reflecting the catabolic stage commonly occuring in advanced HD.83 Despite these results, other studies have failed to detect a significant difference in leptin and ghrelin levels between HD patients and controls.84,85 Further studies of different stages of HD, including prodromal HD, are therefore clearly required to elucidate how and to what extent the neuroendocrine system is affected in HD.
Current and Future Medical Treatment
There is no cure or disease-modifying treatment available for HD.86 However, a variety of symptomatic treatments exist that can have significant beneficial effects on the psychiatric and motor symptoms of the disease.1–3,87 Irritability, depression and anxiety are usually treated with selective serotonin-reuptake inhibitors. Motor symptoms are treated with the monoamine-depleting agent tetrabenazine or neuroleptic agents. The common combination of psychiatric symptoms with depression and/or anxiety, motor symptoms and sleep disturbances often leads to an atypical neuroleptic agent, such as olanzapine, being chosen.2
A larger number of preclinical therapeutic studies and some emerging clinical trials are ongoing for HD.3 Many of the clinical trials are based on promising effects of therapies in animal models of the disease. The possibility to reduce or inactivate the expression of the mutant huntingtin gene with different gene silencing approaches has recently gained significant attention.88–90 Technical concerns remain, as does the choice of which region to target given that not all cells are likely to be reached using this technique. Naturally the striatum, with its severe pathology in human HD, has been the first site of choice for intracranial delivery in animal models. To what extent silencing the mutant huntingtin gene in the hypothalamus and/or the neuroendocrine system will have beneficial effects remains to be tested.
Importantly, hypothalamic dysfunction is implicated in mood disorders, obesity and type 2 diabetes. Intense research and drug development for these conditions is ongoing, with the aim of targeting distinct pathways in the hypothalamus.91,92 The potential of there being common molecular changes between HD and other diseases affecting the hypothalamus opens up exciting possibilities for individuals with HD to benefit from the progress made for these much more common disorders. Likewise, HD with its monogenetic nature may provide a useful system model for similar disorders. Therefore, pre-clinical and clinical studies in HD may possibly facilitate advances that will be beneficial for a larger group of individuals affected by related disorders with hypothalamic dysfunction.
Huntington’s Disease – A Hypothalamic Disorder?
Huntington’s disease has traditionally been viewed as a movement disorder caused by selective striatal pathology. Today, HD is increasingly being recognised as a disorder at the crossroads of neurology, psychiatry, cognitive medicine and genetics. It has distinct but widespread pathology in both the brain and the periphery. It is now clear that one aspect of the disease constitutes hypothalamic dysfunction and pathology. Given the spectrum of non-motor symptoms and signs arising from disturbed functions that are normally, at least in part, regulated by the hypothalamus, it is tempting to speculate that hypothalamic dysfunction plays a role in causing them. Experiments using specific animal models are ongoing to better establish such structure-to-function relationships. Animal studies have been instrumental in getting clinicians to appreciate the non-motor signs in HD and to closer investigate hypothalamic and neuroendocrine changes.46 Translational research will continue to be pivotal for determination of the full extent and clinical significance of hypothalamic and neuroendocrine changes in HD. ■