Stroke induces acute deficits of motion, sensation, cognition, emotion and communication that may occur in isolation or in various combinations depending on the location and size of the infarct lesion. Stroke lesions develop dynamically from the initial lesion corresponding to the area of restricted water diffusion (cytotoxic oedema), which enlarges in most cases into the perilesional hypoperfused region to its final infarct lesion. The neurological deficits typically improve in many patients substantially in the first few weeks after ischaemic stroke. The rate of recovery subsides after the subacute and early chronic phase, but meaningful functional gains are still possible years after a stroke.1 Rehabilitation is a major factor contributing to post-stroke recovery. Notably, patients above 65 years of age can benefit from intensive rehabilitation,2,3 but younger patients typically improve more in areas of mobility, balance, walking and grip strength.4 Rehabilitation of hemiparesis includes passive movements of the limbs to prevent joint contractures, synergistic facilitation of movements of the affected limb and enhancement of active training of the affected limb. Recent rehabilitative strategies are based on findings in systems physiology and functional neuroimaging.5 These strategies aim to prevent learned non-use of the affected limb, improving function by cognitive or imaginative training as well as by non-invasive and invasive cortical stimulation. Surrogate markers of motor impairment and predictors of motor recovery potential may help with determining customised rehabilitative therapies for individual patients and in stratifying patients for experimental rehabilitation studies. Besides behavioural assessments and electrophysiological measures, functional and structural imaging have been proven to be a valuable additional means within this context. Using structural and functional neuroimaging, the human brain and disease-related alterations can be visualised in vivo. Not only can the extent and location of brain lesions be determined, but magnetic resonance imaging (MRI) techniques such as diffusion tensor imaging (DTI) provide additional information about the microstructural status of fibre tracts and the spatial relation between lesions and distinguished tracts. Furthermore, functional imaging allows the study of brain activity related to specific activation states and its changes in relation to adaptation of the brain to a lesion, to deficit compensation and to relearning. This information can be complemented by physiological measures assessing neuronal excitability. In this article we will summarise recent research on determinants of motor recovery such as the extent of damage to major motor fibre tracts. Furthermore, we will describe non-invasive stimulation of perilesional (ipsilesional) and contralesional intact motor cortex in combination with sensorimotor training as a vehicle to enhance motor recovery. These data will be compared with rehabilitative approaches aiming at activation of the mirror neuron system. Additional emphasis will be put on the influence of the somatosensory system for motor recovery after stroke and on secondary changes such as spasticity of the affected limbs.
The Effect of White-matter Damage
Hemispheric brain infarcts typically involve the cerebral cortex. Small cortical lesions may specifically erase a well-defined function that can be localised to this very same area when probed in healthy subjects.6,7 In contrast, larger infarcts usually affect multiple brain systems, which may result in complex neurological syndromes including hemispatial neglect or apraxia. However, involvement of the white matter has not received much attention until recently but was found to be particularly prominent in large cerebral infarcts with and without hemispatial neglect, apraxia and severe hemiparesis.8–11 Notably, it is important to remember that it is not simply the size of the infarct, but preferentially its location that determines the outcome after stroke.12–14
Corticospinal Motor Fibres
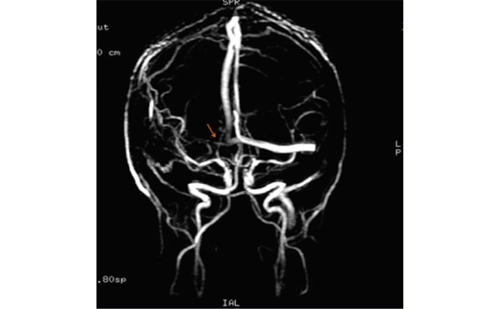
The importance of corticospinal fibres for recovery of motor function after stroke has been demonstrated with imaging and electrophysiological measures.12 Interestingly, clinical and electrophysiological techniques suggested the presence of alternate descending motor fibres (aMF) in addition to the pyramidal tract (PT) since motor evoked potentials (MEP) could still be elicited from the ipsilesional motor cortex15 and selective finger movement was possible16 despite visible damage to the PT. Using DTI imaging as a way of visualising fibre tracts, these alternate motor fibres have recently been visualised (see Figure 1) and their role for motor recovery after stroke has been demonstrated.17 On the basis of evidence from animal work,18 we hypothesise that aMF comprise polysynaptic cortico-reticulo- and cortico-rubro-spinal tracts. The functional integrity of corticospinal motor fibres can be investigated using electrophysiological measures. Transcranial magnetic stimulation (TMS) has been shown to strongly correlate with motor impairment in the acute and subacute phase, whereas its predictive value varied between studies in the chronic stage after stroke.19 In one study, a combination of TMS with DTI-derived parameters proved to be useful in estimating a patient’s potential for recovery when undergoing an intensive motor rehabilitation programme even years after the stroke.1 The most commonly used DTI parameter is fractional anisotropy (FA), which indicates the coherence of aligned fibres and allows inferences of the microstructural status of designated regions of interest or reconstructed tracts.20 FA is calculated from directional diffusivities (axial and radial), which by themselves have been found to reflect the microstructural status of white matter in animal21,22 and human studies.23–25 Axial diffusivity is thought to be an indicator of axonal integrity, whereas radial diffusivity was suggested to primarily reflect (de-)myelination. However, the model of a specific relationship of directional diffusivities with discrete pathological processes is controversial, especially in regions of complex fibre architecture.26 Using diffusivity parameters, fibre degeneration has been revealed in previous studies.27–30 Furthermore, the DTI-based reconstruction of tracts allowed for an evaluation of the topographic relation of a lesion to corticospinal fibres,31–36 the calculation of the overlap between lesion and tracts14 and the quantification of damage to descending motor tracts.1,17,37 The correlations of established motor impairment scores with those DTI-derived measures revealed that DTI can in fact be used as a structural surrogate marker of the amount of damage to corticospinal tracts and, thus, their functional integrity.5
Transcallosal Motor Fibres
In the future, more accurate estimations of recovery potential might be possible when considering not only corticospinal tracts (PT and aMF), but also transcallosal motor fibres. Models of an imbalance in inter-hemispheric interactions after stroke highlight the important role of transcallosal connections for recovery.38 Similarly, functional imaging studies demonstrated an alteration of inter-hemispheric connectivity patterns after stroke,39,40 and experimental non-invasive brain stimulation studies revealed that facilitation of motor recovery can be achieved via upregulation of intact ipsilesional motor cortex and via downregulation of contralesional motor cortex.41 Thus, there is ample evidence for the importance of inter-hemispheric interactions in motor recovery after stroke, although the exact role of contralesional primary and non-primary motor regions remains elusive.42,43 Work in healthy subjects, in which the association of function and microstructure of transcallosal motor connection was demonstrated,44 led to an investigation of those tracts in chronic stroke patients undergoing non-invasive brain stimulation. It could be shown that DTI-derived measures of transcallosal motor-to-motor fibres allowed predictions of therapeutic response to experimental rehabilitation: the more the diffusivity profiles resembled those observed in healthy subjects, the greater a patient’s potential for functional recovery.45
The Role of Perilesional Tissue
As the area of ischaemia typically exceeds the resulting infarct lesion,46–49 an important factor contributing to recovery is the perilesional tissue. The perilesional tissue is supposed to be structurally intact but functionally altered due to transient ischaemia and subsequent reperfusion. Both factors evoke a large number of biochemical, metabolic and immunological processes that evolve sequentially.50 Notably, the binding of flumazenil, a γ-aminobutyric acid (GABA)A receptor antagonist, as measured with positron emission tomography, was found to be reduced in this area in proportion to the initial hypoperfusion as assessed with perfusion computed tomography.51 This suggests loss of inhibitory interneurons in the peri-infarct area and consecutive increased cortical excitability, as demonstrated in TMS studies.52,53 The functionally abnormal perilesional tissue contributes to the clinical deficit, which will affect an activation-related signal: functional MRI (fMRI) performed approximately two days after stroke revealed an area in the ipsilesional postcentral gyrus and posterior cingulate that correlated with motor recovery approximately three months after stroke.54 restoration of hand function three months after stroke was associated with highly lateralised activation of the affected sensorimotor cortex which developed over time.55 Thus, when an impaired function is probed in an activation study, the activation most likely reflects adaptation of the injured brain to the functional deficit owing to spontaneous recovery in the perilesional tissue. The perilesional cortex is anatomically linked to a large number of brain structures that become engaged as a functional network upon the generation of functional activity and in relation to spontaneous recovery. Since the first functional neuroimaging studies in neurological patients with focal brain lesions, it has been well established that there are large-scale changes affecting the contralesional cerebral cortex and subcortical structures in highly structured patterns, which most likely reflects the functional intracerebral connectivity. These functional changes are reminiscent of re-learning, as they represent activation patterns similar to procedural learning and are essentially transient in nature.52,53,56
The Role of the Somatosensory Cortex and the Thalamus
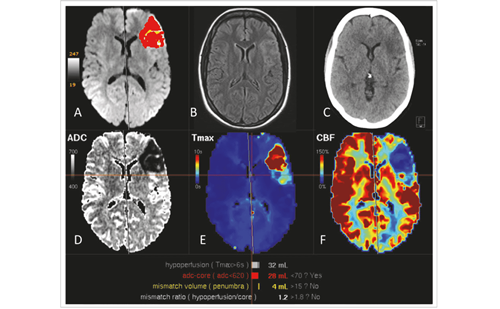
In daily life, intact somatosensation is crucial not only for perception, but also for guidance of action. Accordingly, tactile input, guidance of and activity in somatosensory brain regions have been linked to motor recovery post-stroke.57,58 The severity of touch discrimination impairment experienced post-stroke correlated differentially with brain activity following lesions depending on lesion location in either subcortical or cortical somatosensory regions (see Figure 2). However, notably for subcortical lesions, touch outcome was inversely correlated with brain activity in widespread cortical and subcortical circuits during tactile stimulation of the affected hand.59 In contrast, in patients with cortical lesions there was no correlation between touch discrimination and activation patterns.59 However, activity in the contralesional thalamus was inversely correlated with ipsilesional somatosensory cortex and positively correlated with the contralesional somatosensory cortex.60 It was argued that this would help to redress the imbalance in cortical activity between hemispheres that commonly occurs after stroke and is an important determinant of impairment and recovery.53,61 This could be influenced by interthalamic communication62 or via feedback connections from cortical areas that play an important role in both rapid and more slowly emerging forms of thalamic plasticity.63–65
The Effect of Rehabilitative Training and Underlying Neural Correlates
Rehabilitative training after stroke is known to improve the functionality and to enhance the spectrum of activities of daily living. Functional neuroimaging studies have provided evidence that training has a significant impact on the cerebral activation patterns: Constraint-induced movement therapy, which focuses a patient’s attention to the affected side and involves repetitive training, resulted in improved motor function and enhanced activation in the partially damaged sensorimotor cortex66,67 as well as in other grey-matter structures including the hippocampus.68 Similarly, repetitive training of the affected arm yielded an increase of activation in the sensorimotor cortex related to hand movements, which initially persisted for weeks after training completion and then decreased in magnitude in relation to the functional gains.69 Furthermore, a three-week training in chronic stroke patients using robot-assisted training resulted in improvements of hand motor function which was associated with a greater fMRI signal in sensorimotor cortex related to performance of the movements trained by the robot.70 This increase was task-specific, since it did not occur in relation to non-trained supination/pronation movements with the affected hand and movements of the non-trained hand. Similarly, treadmill training was found to improve walking velocity, which correlated with brain activity in the posterior cerebellum related to movement of the paretic limb.71 Even passive training of wrist movements was reported to be clinically effective and to change the cortical activation,72 although evidence from 3D motor analysis showed that successful hand shaping and grasping of objects did not occur when there was not sufficient volitional control of finger and thumb extensions.73
More recently, cognitive training strategies have been promoted in addition to peripheral sensorimotor activities. It has been assumed that the inferior frontal cortex plays a critical role in motor recovery since there are so-called mirror neurons that become active not only in relation to motor activity but also in response to observation and imagery of the same type of movements. In controlled trials mirror therapy undertaken soon after stroke was found to improve the neurological status immediately after the intervention and at long-term follow-up.74,75 Similarly, mental training was reported to result in better functionality of the upper extremity and in greater gains of activities of daily living than standard physiotherapy.76,77 FMRI revealed that motor imagery activated a widespread network of areas in motor, premotor and parietal cortices in both cerebral hemispheres. Similarly, a daily treatment with observing actions combined with physical training for four weeks resulted in a significant increase in motor functions that lasted for at least eight weeks after training.78 This was associated with a significant overactivation compared with the control group in ventral premotor cortices, superior temporal gyri, the supplementary motor area and supramarginal gyrus related to an object manipulation task. However, it must be mentioned that the capacity to perform motor imagery can be weakened by limb loss or disuse, although the temporal characteristics of motor imagery may be not affected.79
Brain Stimulation as an Add-on to Peripheral Sensorimotor Activities
In the context of experimental rehabilitative therapies, the model of interhemispheric imbalance and the important role of transcallosal connections provide a framework for hypotheses based on two facets: upregulating excitability of intact portions of the ipsilesional motor cortex, and downregulating excitability of the contralesional motor cortex. The contralesional cortex is presumed to be disinhibited due to the lack of an inhibitory influence from the lesional motor cortex while at the same time it exerts an unbalanced inhibitory influence on the lesioned motor region. The downregulation of the contralesional, disinhibited motor regions is presumed to counter an abnormal inhibitory influence on ipsilesional regions (see Figure 3).41,53,80 Pilot studies, using either rapid transcranial magnetic stimulation (rTMS)81–84 or transcranial direct cortical stimulation (tDCS),85–88 have shown that these approaches can improve motor impairment, at least transiently, and that the combination of central stimulation and peripheral sensorimotor activities and training seems to enhance these effects. The efficacy of upregulating ipsilesional motor cortex can be related to its plastic effects on tissue spared by the stroke. As mentioned above, the potential of perilesional tissue for post-stroke recovery has been demonstrated using functional imaging and electrophysiological methods. Accordingly, rTMS yielded therapeutic responses only when at least parts of the motor cortex were spared by the stroke.81 Furthermore, the therapeutic response to anodal tDCS and simultaneous robotic arm therapy was relatively small in patients with extensive hemispheric lesions including the motor cortex.87 It has been argued that, when using large electrodes to target functionally intact perilesional tissue, tDCS can exert its effects not only on the primary motor cortex, but also on adjacent premotor and sensory regions.41 Modulating excitability of such regions, which have previously been shown to play an important role for motor recovery,89,90 may contribute to the efficacy of tDCS.91 Furthermore, it was shown that the enhancing effects of anodal stimulation on the intact portions of the ipsilesional motor cortex85,88 may be potentiated through additional modulation of inter-hemispheric interactions92 via a suppressive effect of cathodal stimulation on the contralesional motor cortex.86 A study in healthy subjects suggested that bihemispheric tDCS (upregulation of affected motor cortex and downregulation of contralateral motor cortex at the same time) produces greater behavioural effects than uni-hemispheric stimulation.93 Accordingly, this novel bihemispheric tDCS therapy with simultaneous physical/occupational therapy for five consecutive days yielded substantial functional improvements that were significantly greater than in a placebo group receiving only physical or occupational therapy.91
The Role of Spasticity
Virtually none of the studies cited above addressed the issue of spasticity, although it is a major sequel of stroke, impairing recovery.94 Spasticity develops within weeks after acute brain lesions, mainly in antigravity muscles such as leg extensors and arm flexors. Spasticity affects movement in terms of velocity and the movement path of limbs. It also requires an extra effort to move the afflicted limbs. One medical treatment option is to inject botulinum toxin (BTX) locally into the motor end plate regions of antigravity muscles to partially paralyse the concerned muscles. Agonists and antagonists immobilised by spasticity prior to injection can then move more freely again. The effect of BTX lasts for about three months until the blocked motor end plates have regenerated entraining a return of spasticity. BTX has been shown to be a safe, effective treatment of upper-limb spasticity caused by stroke or traumatic brain injury.95–97 In a recent study, cyclic ergometer training prolonged the antispastic effect of BTX injection and yielded an increased range of motion of the paretic arm. In patients with residual motor function the decrease of spasticity due to combined cyclic ergometer training and BTX injection into forearm muscles was paralleled by an increase of fMRI activity in relation to passive arm movements in the dorsomedial portion of the sensorimotor cortex in the lesioned hemisphere and in the secondary somatosensory area of the non-lesioned hemisphere (see Figure 4). In contrast, there was no training-induced increase of fMRI activity with passive arm movements in completely paralysed patients, suggesting that in these patients both the efferent motor fibres and the afferent somatosensory fibres were severely damaged.98
Conclusion
High-resolution structural and functional brain imaging (including DTI) and TMS as diagnostic tools to assess motor evoked potentials constitute a powerful combination to explore the normal structure and function of the motor system as well as alterations of motor circuits caused by a stroke. Parameters derived from these modalities of systems physiology can be used as surrogate markers of the motor system’s functional integrity. Clinicians can potentially rely on this physiological information to determine which rehabilitation strategies are most appropriate for individual patients. Further, future developments in rehabilitation may employ this physiological information for designing innovative rehabilitation approaches and for predicting the therapeutic response to such interventions. ■